
研究紹介
当研究室では、動物細胞のシグナル伝達系とその異常によって引き起こされる病態に関連した研究を行っています。生化学、分子生物学、細胞生物学の研究手法を駆使し、生命現象の新規の調節メカニズムの解明を目指しています。さらに、そこから得られる知見を基盤とした応用研究への発展の可能性も追求します。
(1)細胞内シグナル伝達系
細胞は常に外界の環境の変化を感知して、それに適切に応答する能力を有している。動物の生体内においては、細胞は、体液中に存在する分子や隣接する細胞との相互作用を通じて、外界からの様々な刺激を受容している。細胞外の(水溶性)リガンドは、細胞膜上に存在する特異的な受容体に結合して、そのコンフォメーション変化(活性化)を誘導する。活性化された受容体は、その細胞内領域において、種々のシグナル伝達分子と会合してシグナル伝達複合体を形成し、下流へとシグナルを伝達する。シグナルは、第二メッセンジャーと呼ばれる低分子化合物の濃度変化やシグナル伝達蛋白質間の相互作用を通じて伝達される。最終的に、核における遺伝子発現や細胞骨格系などが制御され、様々な細胞応答を引き起こす。
(2)GTP結合蛋白質
細胞内シグナル伝達系で機能するGTP結合蛋白質は、三量体として機能する「G蛋白質」と単量体として機能する「低分子量GTP結合蛋白質」に大別される。GTP結合蛋白質は、結合しているGTPをGDPと無機リン酸に加水分解する酵素活性をもつことから、GTPアーゼ(GTPase)と呼ばれることもある。また、GTP結合蛋白質の略称としてG蛋白質という表現が用いられる場合も多い。GTP結合蛋白質は、GTPまたはGDPと非共有結合し、細胞外刺激に応答して誘導されるGTP型(シグナル開)とGDP型(シグナル閉)との間での変換を介して、シグナルの開閉を行う分子スイッチとして機能している。GDPとGTPとの交換反応を制御する蛋白質は、グアニンヌクレオチド交換因子(GEF; guanine nucleotide exchange factor)と呼ばれる。一方、GTP加水分解活性(GTPアーゼ活性)を制御する蛋白質は、GTPアーゼ活性化蛋白質(GAP; GTPase-activating protein)と呼ばれる。通常、シグナル伝達に対して、前者は正に後者は負に作用している。
(3)インスリンの作用と糖取り込みのシグナル伝達系
インスリンは膵島β細胞から分泌されるホルモンで、肝臓、骨格筋、脂肪細胞、脳などの臓器を標的として種々の生理反応を引き起こし、血糖値を低下させる作用をもつ。骨格筋や脂肪細胞においては、インスリンは血中のグルコースを細胞内に取り込ませる。この際、インスリン受容体を介して活性化される細胞内シグナル伝達系は、糖輸送担体GLUT4のGLUT4貯蔵小胞(GSV; GLUT4 storage vesicle)から細胞膜表面への移行(小胞輸送および細胞膜との結合・融合)を誘導する。細胞膜に局在するGLUT4を介する促進拡散により、グルコースが細胞内に取り込まれることが知られている。当研究室では、骨格筋におけるインスリンおよびその他の外界からの刺激に応答した糖取り込みのメカニズムの解明を進めている。とくに、低分子量GTP結合蛋白質Rac1がこのシグナル伝達系で重要な役割を果たしていることを見出し、その調節機構の解明を進めている。培養細胞株および遺伝子操作マウスを用いた解析(遺伝子導入、遺伝子ノックダウン、蛍光抗体法によるGTUT4の細胞膜への移行の観察など)を進めている。
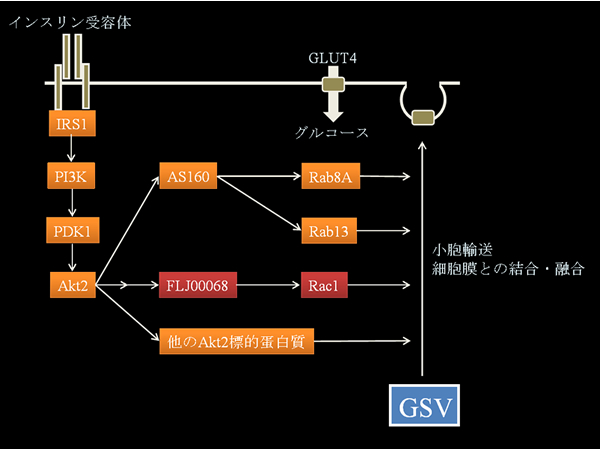
図の説明:骨格筋におけるインスリン依存性糖取り込み誘導シグナル伝達系。インスリン受容体からのシグナルは、IRS1を介して、PI3K、PDK1、Akt2からなるキナーゼカスケードに伝達される。Akt2は、種々の標的蛋白質をリン酸化することにより、下流にシグナルを伝達する。AS160は、Rab8AおよびRab13に対するGAPであり、Akt2によるリン酸化は、GAP活性を負に制御し、Rab蛋白質の活性化を引き起こす。当研究室では、Akt2の下流で、GEFの一種であるFLJ00068を介して、RhoファミリーGTP結合蛋白質の一種Rac1がGLUT4の細胞膜への移行を調節していることを見出した。AS160以外にも、GLUT4の細胞膜移行の制御に関与するAkt2の基質が存在している。
(4)中心体複製の制御機構
中心体は、直角に結合した2つの中心小体とそれを取り巻く中心小体周辺物質からなる細胞小器官(微小管形成中心)であり、細胞分裂時には紡錘体の極として重要な役割を担っている。中心体は、細胞周期の進行に伴い、半保存的に複製されるが、染色体の複製や細胞質分裂と同調するよう、厳密な制御を受ける必要がある。実際、中心体の複製サイクルにおいては、多数の因子の関与が報告されており、各因子の役割の詳細も解明されつつある。中心体複製の制御からの逸脱は、分裂装置の多極化と染色体の分配の異常を引き起こすことから、細胞の異数性の原因の1つと考えられている。当研究室では、グアニンヌクレオチド交換因子ARHGEF10が、低分子量GTP結合蛋白質RhoAの活性化を介して、中心体の過剰な複製を抑制するメカニズムに焦点を当てて、共焦点レーザー走査型顕微鏡などを駆使して、研究を進めている。
![]()